Using genetics to conserve life history strategies in California steelhead (Oncorhynchus mykiss)2/4/2021
This blog post was originally published by the AGA Blog in February 2021.  Credit: Timothy Knepp – U.S. Fish and Wildlife Service (National Digital Library of the USFWS) By Alexandra L. DeCandia Steelhead (Oncorhynchus mykiss) inhabit cultural and ecological niches in northern California and the Pacific Northwest. Culturally, steelhead provide an important food source to indigenous communities and represent abundance, wealth, prosperity, and determination in traditional art and religious ceremonies (Squamish Lil’Wat Cultural Center). Ecologically, migratory steelhead transport key nutrients between marine and freshwater environments (and the organisms that inhabit them) and serve as indicator species for ecosystem health. Diverse life history strategies adopted by steelhead are critical to the fulfillment of these niches. Perhaps most emblematic concerns migration. Steelheads are classified into two primary groups: residents and migrants. Resident individuals inhabit streams, rivers, lakes, and reservoirs throughout their lives, whereas anadromous steelhead migrate between marine and freshwaters for feeding and spawning. Within the migrating group, there are additional strategies regarding the timing of migration from marine to freshwaters (termed “run-timing”; Figure 1). Summer-run steelhead migrate to freshwaters in late spring/early summer to mature in the fall and spawn in winter. In contrast, winter-run steelhead wait until late fall/winter to migrate into freshwaters for spawning. 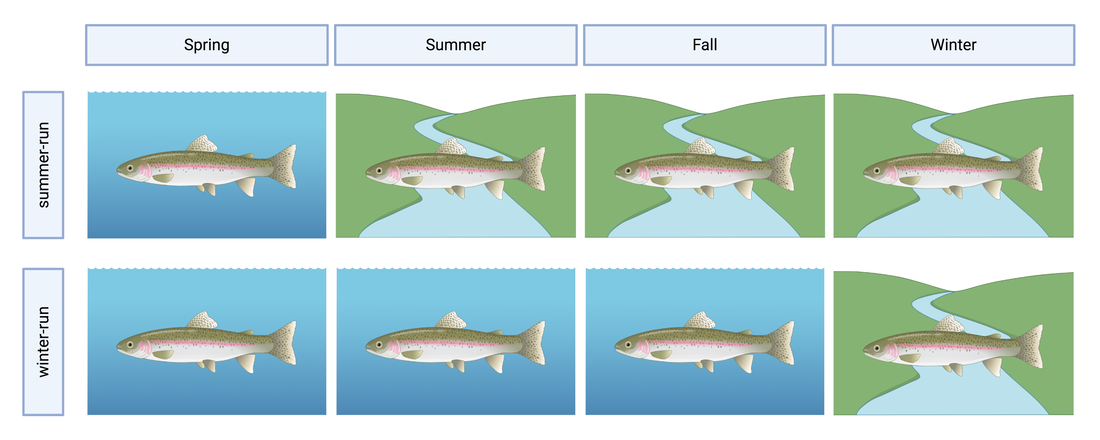 Figure 1. Simple schematic of summer-run and winter-run migration timing. Figure created by A. DeCandia with BioRender. Although once abundant throughout the region, steelhead have experienced dramatic declines due to anthropogenic activities. Stressors include dams, diversions, development, pollution, run-off, harvest, thermal stress, and reduced stream flow that render certain routes impassable. In addition to causing the loss of individuals, these threats jeopardize the complex life histories exhibited by steelhead in the region. Summer-run steelhead are particularly vulnerable and at risk of local extirpation. In their recent study, Samantha Kannry et al. (2020) examined the distribution of life history strategies in the southernmost river system with summer-run steelhead: the Eel River in northern California (Figure 2). More specifically, the authors used genetic techniques to examine population structure, demography, and ecology throughout the Eel River watershed. Their dataset consisted of genome-wide loci genotyped through Rapture sequencing, as well as markers previously associated with migration (OMY5) and run-timing (GREB1L). Their sample set included more than two thousand steelheads sampled for this and a previous study in the Eel River watershed. 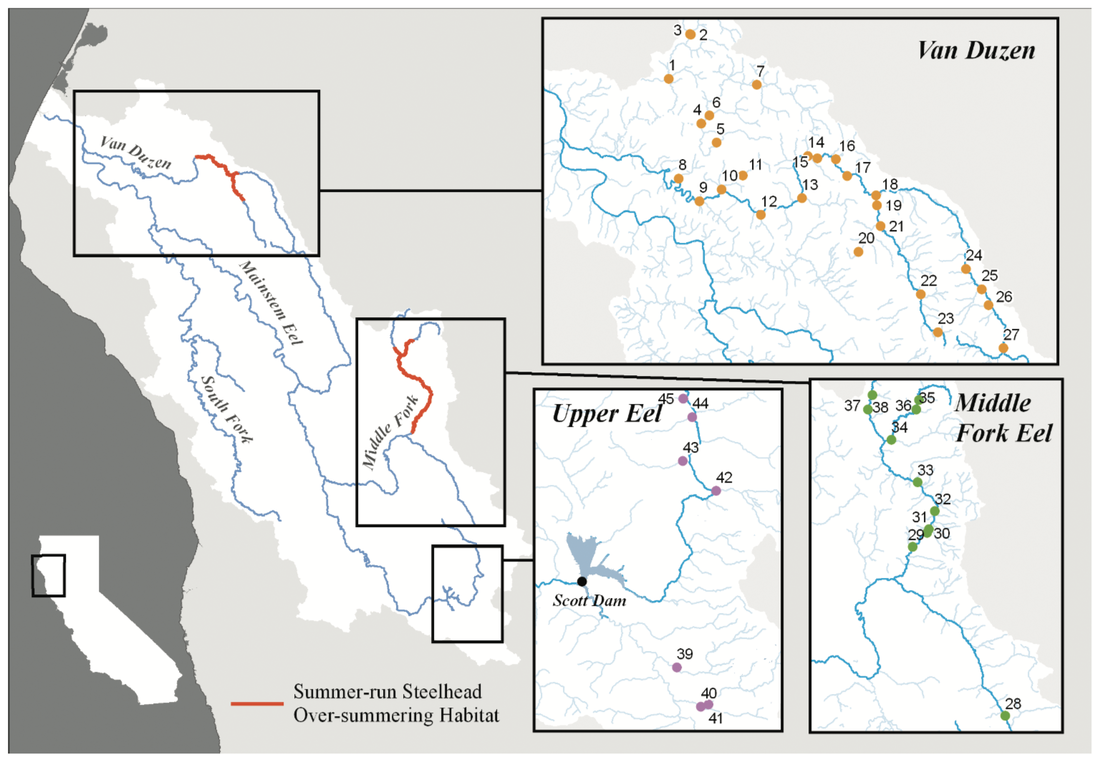 Figure 2. Map of study area (Figure 1 in Kannry et al. 2020). Principal component analysis revealed three genetic clusters within the watershed. Interestingly, steelhead primarily clustered by sampling basin rather than life history phenotype. Summer-run and winter-run individuals sharing the same sampling basin (e.g., Van Duzen) therefore clustered more closely together than summer-run steelheads sampled across basins. This result supported the presence of two independent populations of summer-run steelhead in the Van Duzen and Middle Fork Eel Rivers. Critically, this overturned the previous assumption that all summer-run steelhead derived from the same subpopulation. Kannry et al. next considered potential barriers to steelhead passage found throughout the watershed. Through examination of migration allele frequencies, they reported that Eaton Falls does not present a complete barrier to anadromous steelhead, as previously thought. This result was supported by estimates of population differentiation across no barrier (mean pairwise FST=0.059), the partial Eaton Falls barrier (0.094), and the confirmed South Fork barrier (0.219). Within migrating steelhead, they also identified barriers between winter-run and summer-run steelhead in both the Van Duzen (Salmon Falls barrier) and Middle Fork Eel (Osbourne Roughs barrier) rivers, suggesting habitat segregation between the two run-timing strategies. This result found its extreme in the South Fork Eel River, where no summer-run alleles were identified, thus indicating the absence of this strategy from this part of the watershed. The final barrier considered by the authors was Scotts Dam, a hydropower facility that has diverted water into the nearby Russian River watershed for roughly 100 years. Constructed without a fish passage, Scotts Dam presents a formidable barrier to anadromous steelhead. As this dam is up for relicensing in 2022, Kannry et al. considered whether the migration phenotype could be recovered above the dam. Through population genetic surveys, the authors identified both anadromous and summer-run alleles in the resident steelhead population sampled above Scotts Dam. This suggests that steelheads may be able to naturally recover this important strategy without human intervention, should the dam be removed. In summary, the authors identified distinct population segments, mixed and distinct resident and migrant waters, candidate areas for habitat restoration, and potential source populations for the natural recovery of life history strategies. These insights can directly inform the conservation and management of steelhead in the Eel River watershed, particularly regarding the 2022 Scotts Dam relicensing. In addition to these implications, the analytical approach described by Kannry et al. provides a roadmap for conducting similar analyses in other river systems in northern California and the Pacific Northwest. By better understanding the ecology and distribution of life history strategies throughout the region, the authors demonstrate that diversity can be preserved. Given steelhead’s significance, these analyses and management recommendations may enable steelhead to occupy important cultural and ecological niches indefinitely. Comments are closed.
|
